May 16, 2024
Frozen brain tissue brought back to life in major breakthrough
Posted by Dan Breeden in categories: innovation, neuroscience

*BREAKTHROUGH!!*
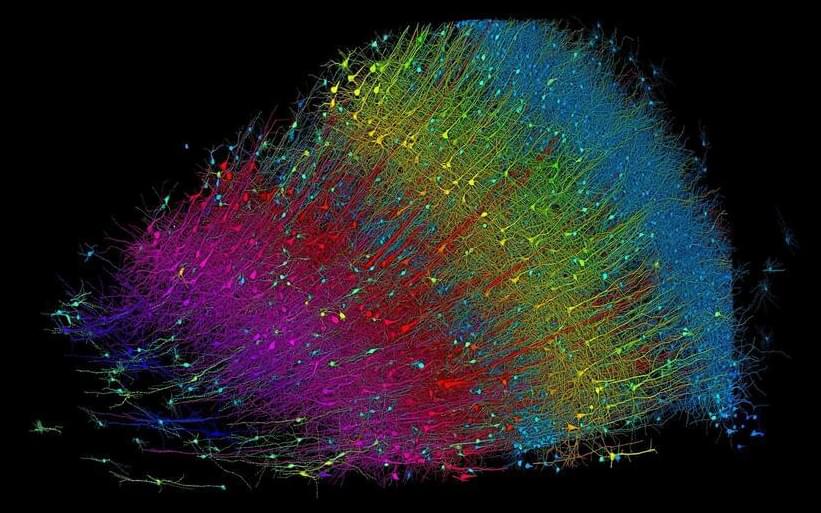
Scientists may one day be able to freeze brains and bring them back to life following a major breakthrough in cryogenics.
Researchers in China have successfully frozen and thawed human brain tissue, after which it regained normal function.
Continue reading “Frozen brain tissue brought back to life in major breakthrough” »